









Master Universitari
Master universitari di II livello in ossigeno ozono terapia Sioot con valore lagale.


Il primo protocollo scientifico di ossigeno ozono terapia al mondo. Prevenzione e cura del Covid19 e del post Covid19.
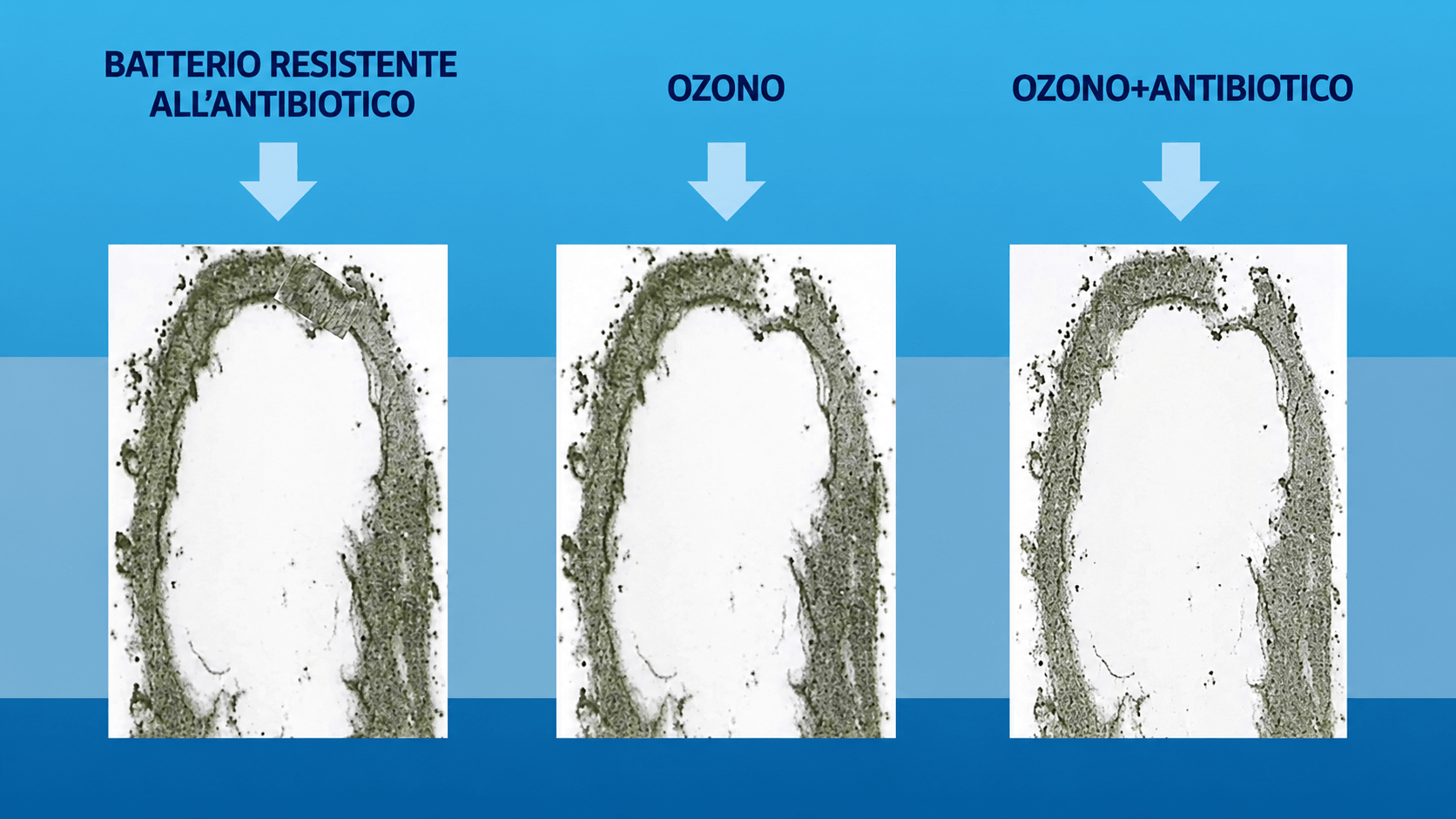
I meccanismi d’azione dell’ozonoterapia nel trattamento dell’infezione da virus SARS.





L’OZONOTERAPIA SIOOT È LA SOLUZIONE
Perché l’ozono?
Bibliografia disponibile a richiesta – oltre 1400 titoli
Visualizza e scarica le domande di adesione SIOOT e ASOO
Master universitari di II livello in ossigeno ozono terapia Sioot con valore lagale.